






総論 獲得免疫・免疫遺伝
総論 獲得免疫・免疫遺伝
熱研・免疫遺伝学分野 平山謙二
感染症は熱帯病の中心
伝染病の発生には環境の要因が重要で、特に熱帯地域は経済的社会的な遅れから劣悪な衛生状況に陥やすく、種々の感染症が蔓延することとなる。表1に示すようにアフリカでの乳幼児死亡の原因のトップは、マラリアでその他、肺炎や下痢など病原微生物による死亡は、現在最も問題となっている。
感染性微生物として、重要なのはウイルスから大型寄生虫まで多様なものがあるが、ウイルスでは、黄熱、デング熱、肝炎、ハンタウイルス、細菌では、結核、らい、サルモネラ、コレラ、病原大腸菌、赤痢、髄膜炎など、寄生虫では、マラリア、赤痢アメーバ、リーシュマニア、トリパノソーマ、フィラリア、住血吸虫、鉤虫などである(図1)。
上記のような感染症については、これから各論で講義あるいは実習を通して、それぞれの感染経路や臨床像、病原体の様子について理解してもらうことになっている。
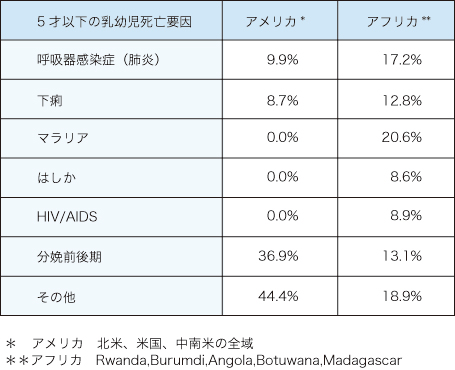
表1. 乳幼児死亡要因 2000年 WHO, CHO
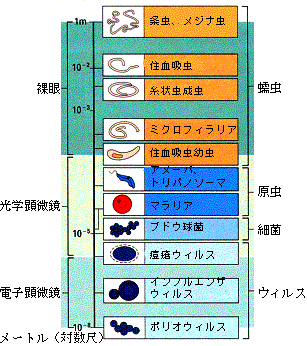
図1. 各種寄生体の大きさの比較
生体防御反応
これらの感染性微生物は感染形式、組織分布などは異なるがいずれも外来の異物として生体内に侵入するので、一様に生体防御反応と遭遇することになる。生体防御反応とは、ヒトが、有害な外来異物から身を守るすべての生体反応の総称である。熱帯地域で自分の身を守るためには、この全体としての生体防御について常に考える必要がある。生体防御にはいくつかのレベルがあるが、大きく分けると神経系によるものと免疫系によるものである。神経系による生体防御は五感と脳の統合野を使った防御で、たとえば、痛いものには触れないし、臭いものは食べないし、危ないと知っているものには近づかないといった反応である。咳やくしゃみ、嘔吐、下痢なども主に神経による反応であろう。AIDSの流行地で性的な接触をしない、コレラを避けるために生のカキを食べない、マラリアの媒介蚊に刺されないように夜出歩かないなどは、高度な神経による防御反応である。
強調しておきたいのはこの神経レベルでの防御反応、特に脳のレベルでの反応は熱帯病では最も重要だということである。神経系の防御をかいくぐると、次に免疫系のお出迎えである。免疫というと「2度がかりなし」という獲得免疫が古来から知られているが、この30年来の免疫学の進歩により、免疫系のシステムとしての全容がほぼ明らかになりつつある。すでに自然免疫の項で触れられたことと思うが、この免疫システムは2つに分けて考えることが可能である。1つは自然免疫natural immunityもう1つは獲得免疫aquired immunity である。いずれも外来異物が不幸にして侵入してきたときにこれを排除するように作られた体のしくみである(図2)。自然免疫と獲得免疫の最も大きな違いは免疫学的な記憶があるかどうか、すなわち2回目に出会った異物に対しての反応性の効率が同じか増大するかという違いである。この免疫記憶という概念はきわめて文学的で科学としての文脈に乗らないものであるが、それが徐々に科学的に解析され、分子レベルで明らかになりつつある。自然免疫といえどもまったく記憶がないわけではなく、種あるいは生物としての古来の記憶(たとえば、細胞の莢膜のある糖鎖を認識するなどはこの例であるが)によって反応している、ということもできる。まとめると種あるいは生物としての記憶による免疫(生体防御)が自然免疫、個体の記憶による免疫が獲得免疫という風に考えることもできるのである。
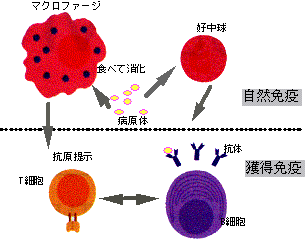
図2. 自然免疫と獲得免疫
自然免疫
自然免疫は主に外界との境目で働くことが多いが、皮膚、消化管肺粘膜などで特に発達している。皮膚自体は皮脂腺などによる防御能を持っているが、肺粘膜などは繊毛が生えており、異物を外界へと送り出す作用をしている。消化管粘膜は広げるとテニスコート一面分あるそうだが、この巨大な外界との接触面にも自然免疫が発達している。その最前線は粘液や蠕動運動である。一度このバリアーを突破すると次に強力な免疫細胞や免疫分子が出動してくる。それが、貧食細胞と補体である。貧食細胞であるマクロファージや顆粒球は様々な異物を個体としての記憶なしに認識し、細胞内にとりこみ消化してしまう。また補体は普段は非活性の安全なタンパク質のセットであるが、一度、引き金が引かれる(すなわち、タンパク質が活性型に変化する)と恐ろしい毒性物質となって外来異物に襲いかかり、殺滅してしまう。このシステムは、体中にはりめぐらされた盗難者検知赤外線ビームのようなもので触れたら最後大騒ぎを引き起こす仕組みになっている。これ以外にもナチュラルキラー細胞(NK細胞)という特殊なリンパ球(白血球の一種)が知られており、主にガン化した細胞を殺して生体を維持する働きをしている(生体監視機構)と考えられている。
獲得免疫
元々2度がかりなしという概念がいつ頃できてきたのかは不明だが、病原体という概念の発生によって気づかれたものと思われる。乳しぼりの歌にあるように牛痘にかかった者が、痘瘡に抵抗性になることは昔から知られていた。ジェンナーは1796年牛痘の膿をナイフで少年に接種し、見事に感染を防御することを示した(動物実験をやらずこんなに恐ろしい実験ができたのも時代のせいだろうか)。ワクチンの発見である。
抗体あるいはB細胞のはたらき
その後、この防御能を担うのが血液中の血清成分に含まれることが北里柴三郎により示され、抗体と名づけられた(図3)。血清療法の始まりである。外来の異物は何千通りも種類があるのに抗体はどうやってこれを見分けるのか。これを解明したのが利根川進らのグループである。抗体を作るのが特殊な白血球(B細胞)であることは知られていたが、このB細胞の腫瘍であるミエローマ(B細胞腫)の抗体遺伝子が普通の体細胞の抗体遺伝子から変わっていることを発見したのである。これは、それまでの遺伝子は不変であるという常識を一変するもので、これにより、多種多様な抗体分子が作られるメカニズムが明らかになった(図4)。
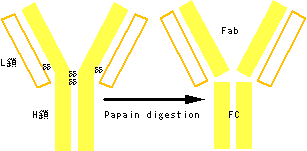
図3. 抗体の構造
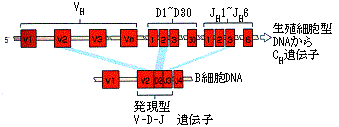
図4. ヒトのVDJ遺伝子組み換え
抗体はどのようにして作られるのか
抗体はB細胞とよばれる特殊な白血球(リンパ球)により作られるが、一個のB細胞は一種類の抗体を作ることが知られている。これは、このB細胞が先に述べたようにすでに抗体遺伝子を変化させて、特有の抗体を作るように分化してしまっているからである(図5)。また、この際一個の細胞には父方と母方の2個の抗体遺伝子を持っているが、実際に作られるのは、その一方だけである。こうして、多様な(109種といわれている)抗体遺伝子を持ったB細胞前駆体が、リンパ節やパイエル板などの免疫組織(トリではBursa)で分化し、流血中やリンパ節で外来異物由来のタンパク質や糖といった抗原との相遇を待っている。皮膚や消化管、呼吸器粘膜などから侵入した病原体は自然免疫をのがれると増殖し、大量の抗原を放出するが、これを補食した貪食細胞の一部は血流あるいはリンパ流に乗りこれに反応する抗体を有するB細胞は抗原と反応し、所属リンパ節へ向かう。ここでB細胞エリア、胚中心と言われる場所に定着し(図6)、抗体産生細胞(プラズマ細胞)に分化し、抗体を産生する。抗原は樹状細胞によって保持され、後述するT細胞の補助で、さらに、記憶B細胞の分化増殖がおこり、いわゆるバーネットのクローン選択が完了する(図7)。
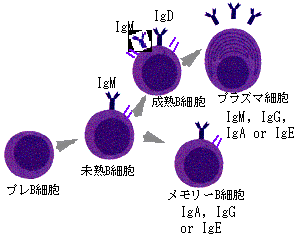
図5. B細胞の分化
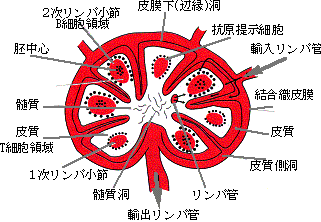
図6. リンパ節の模式図
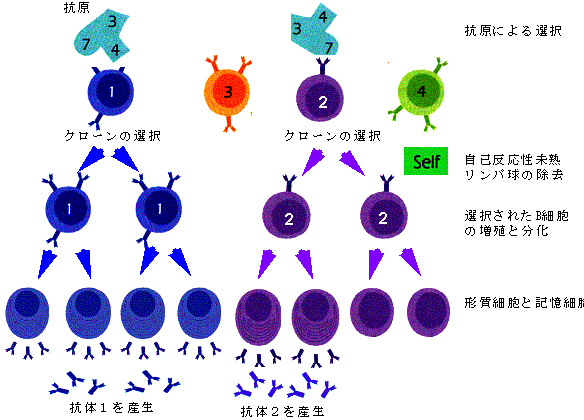
図7. B細胞クローンの選択
T細胞のはたらき
獲得免疫の記憶は抗体やB細胞だけではなく、T細胞という別の種類の白血球(リンパ球)によっても担われていることがその後明らかとなった。T細胞のTは胸腺(Thymus)の頭文字からとったものであるが、胸腺で分化した細胞という意味である。T細胞は、B細胞と同様、細胞表面に一種類の完全な抗原受容体、T細胞レセプターを表現している(図8、10)。
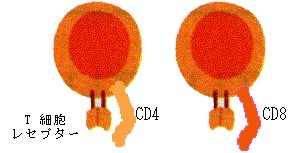
図8. T細胞の構造
この受容体は抗体とは若干趣きが異なり、抗原と自己の抗原提示分子(MHC;Major Histo-compatibility Complex)(図9)とくっつく構造をしている。このT細胞は胸腺を通りぬけることで、成熟したT細胞へと分化し(図11)、流血あるいはリンパ節のT細胞エリアに存在して抗原刺激を待つこととなる。

図9. MHCの複合体の構造
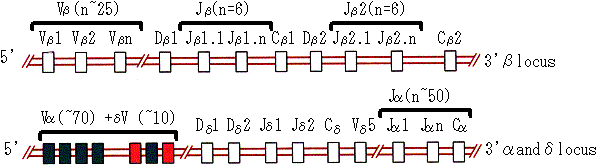
図10. T細胞受容体遺伝子
T細胞がB細胞と異なるのはその抗原受容体であるT細胞受容体が抗原とMHCの複合体と反応することで、そのためこの刺激には、特殊な抗原提示細胞が必要となるAPCとしては、B細胞、マクロファージ、樹状細胞、ランゲルハンス細胞などがあるが、これらの細胞は抗原を細胞内にとりこんだ後、この抗原(とくにタンパク質)をリソソームという袋の中で消化し、MHC(ヒトではHLAと呼ばれる:後述)と結合させて、10個ほどのアミノ酸よりなるペプチドとしてこれを再び細胞表面に提示することとなる(図12)。
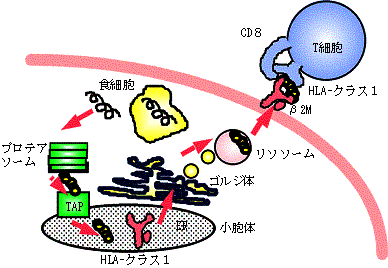
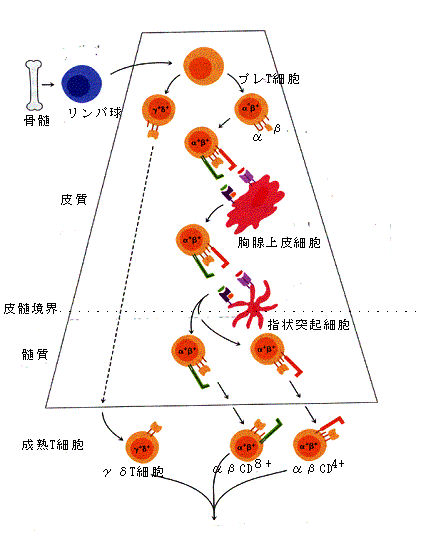
図11. 胸腺におけるT細胞の分化
図12. 抗原のプロセッシングとHLAによる抗原提示
こうして提示された抗原MHC複合体により、これと適合するT細胞受容体を持ったT細胞は活性化し、主にリンパ節内で増殖し、実際の免疫応答の場に戻っていったり、記憶細胞としてリンパ節にとどまったりしている。B細胞と同様にT細胞も骨髄の前駆細胞から分化し、多様なT細胞受容体を持った細胞を次々と作っているが、分化の主要な場とされる胸腺は青年期以降退縮していき、成人では痕跡となってしまう。胸腺を通らないT細胞はおそらく分化できずそのままアポトーシスを起こして死んでしまうと思われる。こうしてまわりくどいやり方(実は意味がある。MHCが出現
してくるのは両生類からであるが、ある説ではウイルスなどの細胞内感染した自己細胞を殺すために、どうしても自己+抗原を認識する必要が生じたとされている。わかる?)で抗原刺激を受けたT細胞は、活性化して増殖し、各種のサイトカインを出して、B細胞やマクロファージを活性化したり(ヘルパーT細胞)、直接効果分子(パーフォリンやグランザイム)を出して標的細胞を殺滅したり(キラーT細胞)する(図13)。
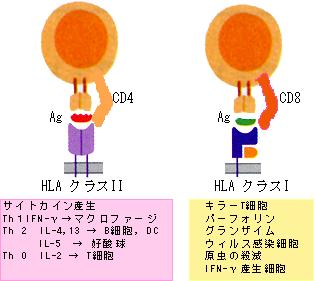
図13. エフェクターT細胞の働き
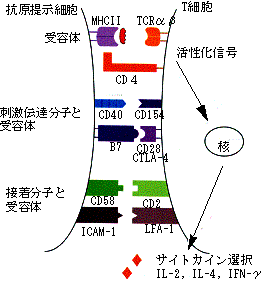
図14. 細胞表面分子とT細胞の活性化
こうして活性化されたT細胞には、上記のように様々な機能があることが知られている。どういう機能をもったT細胞に分化するかは、APCからT細胞への刺激の際の、各種リセプター、リガンド及びケモカインやサイトカインの共刺激などによって決定されると考えられている(図14)。
貪食細胞、抗原提示細胞について
マクロファージは、自然免疫の主要なプレーヤーとして重要だが、貧食細胞系としては各組織に種々の特異な細胞があり、各組織で特有の働きをしていると考えられている。皮膚には、ランゲルハンス細胞という樹状細胞が一面にはりめぐらされており経皮侵入した病原体は、この細胞により補促され、リンパ節へと運ばれる。流血中にも分化あるいは未分化の樹状細胞(DC)が存在し、ある種の刺激、たとえばサイトカイン(GM,CSF,IL-4)により分化し、非常に効率の高い抗原提示細胞となる。また、肝臓の類洞にも特殊な免疫組織が存在し、クッパー細胞という貧食細胞が抗原提示を行う。脳や中枢神経系では星細胞が、間葉系由来の貧食細胞として知られているが、免疫系における機能についてはまだ不明の点が多い。
エフェクター細胞について
での話は、免疫記憶に関与する細胞及びシステムについてであったが理解してもらえたであろうか。進化の過程では、まず貧食細胞や補体といった自然免疫が発生し、さらに抗原を中和したり、貧食しやすくする抗体が現れ、ウイルス感染細胞を排除するために自己MHCと抗原を認識するT細胞が出てきて、ほぼ現在のヒト免疫系が完成したと考えられている。進化の度に免疫システムはより繊細でより強力になって来た。この講義の最後に最終的な免疫効果を及ぼすエフェクター細胞という概念で獲得免疫を整理したい(表2)。
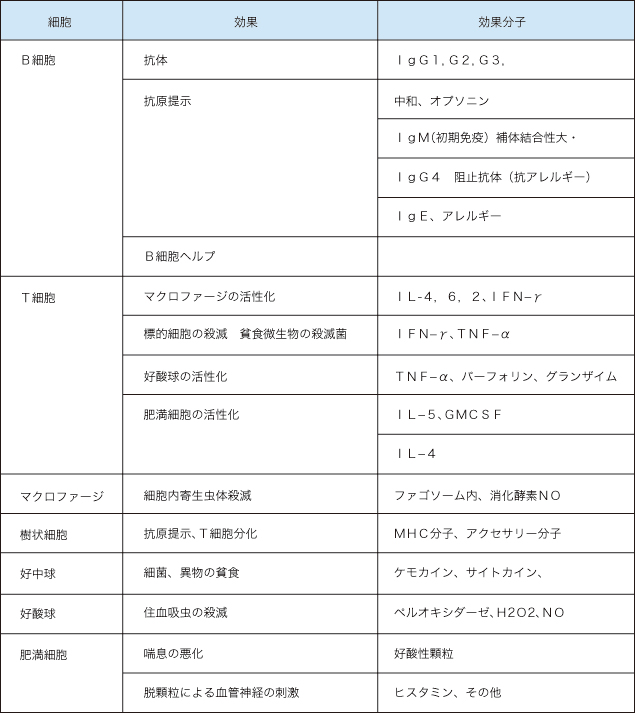
表2. エフェクター細胞による免疫効果
効果分子として挙げたものをすべて詳述することはできないが大別すると抗体、サイトカイン、ケミカルメディエーター、活性酸素、酸化窒素となる。すべて効果分子として重要であるが主だったものについて解説する。抗体分子の機能について特に詳述する必要はないが、1gG4以外の1gGについて特に機能的な定義はない。中和やオプソニン作用、ADCCなどである。1gG4は、1gEと共にアレルギー反応と関係し、Th2細胞のヘルプで産生される。住血吸虫などでは、1gEの効果を阻害する働きがあるとされる。1gAは消化管への分泌抗体で消化管免疫の主力である。1gEは特に肥満細胞上のFcεRと結合して抗原による架橋を起こすことで脱顆粒を引き起こす。サイトカインのうちT細胞が産生するIL-2,4,5,6, IFN-γ, TNF-αはいずれもサイトカインによる効果細胞の活性化に重要である。IL-4はTh2細胞の分化増殖に必須である。Th1の分化にはIL-12(主にマクロファージ系が産生)が必須である。IL-5は好酸球の増殖に重要である。IL-6はB細胞の分化や、肝細胞の刺激分子として多機能である。
IFN-γ、TNF-αはマクロファージの活性化に重要で特にIFN-γの産生が阻害されると感受性微生物を防御できなくなることという多くの報告がある。TNF-α はマクロファージからも産生され、炎症の増悪と関係している。パーフォリンやグランザイムは細胞障害性CD8T細胞の効果分子で前者は補体と同様膜上に穴をあけ後者はその穴から侵入して細胞の核内の遺伝子をズタズタにしてアポートシスをおこす。TNF-α系は、アポートシスを引き起こす因子として重要で、TNF Receptorを介したアポートシス誘導シグナル伝達系が詳しく研究されている。ファゴゾーム内の消化酵素はリソソームとよばれ、システインプロテアーゼを始めとして各種の蛋白分解酵素が混在している。活性酸素O—やH2O2—、NO一酸化窒素などには直接病原体を障害する作用があって、主にマクロファージや好中球で大量に作られる。肥満細胞の顆粒内にはアレルギー反応を引き起こすケミカルメディエーターとして、ヒスタミンなどが存在している。
